

Journal of Translational Genetics and Genomics





Andrea M. Gross , ... Brigitte C. Widemann
, ... Brigitte C. Widemann
, ... Brigitte C. Widemann
Views: Downloads:
Views: Downloads:
Ahmed Kamal, ... Eslam M. El-Nahrery
, ... Eslam M. El-Nahrery
Views: Downloads:
Roshni Bibi, Koustav Sarkar
, Koustav Sarkar
Views: Downloads:
Views: Downloads:
Views: Downloads:
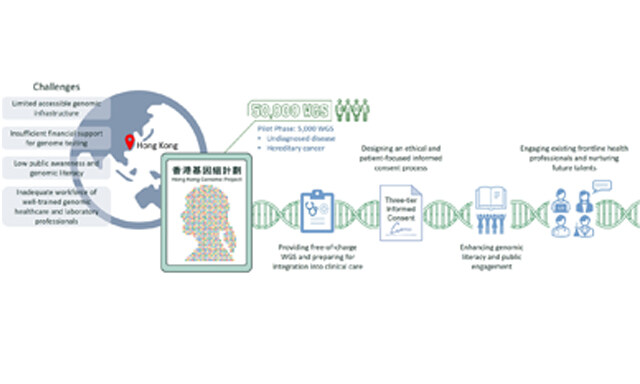
Annie T. W. Chu, ... Brian H. Y. Chung
Views: Downloads:
Amy Hunter, ... Kate Thomson
, ... Kate Thomson
Views: Downloads:









Data
894
Authors
399
Reviewers
2017
Published Since
483,385
Article Views
131,483
Article Downloads
For Reviewers
For Readers
Add your e-mail address to receive forthcoming Issues of this journal:
Themed Collections
Volume 8, Issue 1 (in progress)
Volume 7, Issue 4
Volume 7, Issue 3
Volume 7, Issue 2
Volume 7, Issue 1
Volume 6, Issue 4
Volume 6, Issue 3
Volume 6, Issue 2
Volume 6, Issue 1
Volume 5, Issue 4
Volume 5, Issue 3
Volume 5, Issue 2
Volume 5, Issue 1
Volume 4, Issue 4
Volume 4, Issue 3
Volume 4, Issue 2
Volume 4, Issue 1
Volume 3
Volume 2
Volume 1
Data
894
Authors
399
Reviewers
2017
Published Since
483,385
Article Views
131,483
Article Downloads


